PlantVital®5000
Измерительная
система PlantVital®5000
инструмент
для
объективного
доказательства
вегетационного
состояния растительности
1.
Введение
Измерительная
система PlantVital®5000
позволяет
определять
жизнеспособность
хлорофилл «а»
содержащих
видов на
основе
выделяющегося
в процессе
фотосинтеза
кислорода.
Описание
технических
данных
измерительной
системы PlantVital®5000
представлено
в
Информационных
письмах 01/04 и 02/04.
Философия
измерительной
системы и стратегия
измерения
подробно
описаны в
Информационном
письме 03/04 и
Аппликационном
письме ¼,
соответственно.
Ниже приведены
примеры
важного
вклада
нового метода
биоиндикации
при
установлении
непрерывного
контроля за
развитием
вегетации растительности
в
промышленно-напряженных,
сельскохозяйственных,
а также
заповедных
зонах. Таким
образом,
этот
метод может
служить основополагающим
для
заблаговременного
проведения
мероприятий,
предотвращающих
опасность
для
окружающей
среды в случае
обнаружения
отклонений
вегетации в жизнедеятельности
растений.
Городские
растения
имеют
большое
санитарно-гигиеническое
и
эстетическое
значение для
человека. Они
украшают
город, помогают
удалять
многие
вредные
вещества из
городской
среды,
создают
особые
микроклиматические
условия,
уменьшая
скорость
ветра, снижая
шум, затеняя
территории.
Разумеется,
городская
растительность
нуждается в
защите и
уходе, т.к.
урбанизация
оказывает
большое
влияние на
состояние
растений.
Химическое
загрязнение,
присутствие
пыли в воздухе,
уплотнение и
загрязнение
почв являются
основными факторами
негативного
влияния
городских
территорий
на
растительность.
При этом изменяются
индивидуальные
морфологические
и
физиологические
параметры
растений, нарушается
их генотип,
затормаживается
рост и
развитие
растений,
снижается их
устойчивость
к таким
факторам, как
засуха,
холод,
вредители.
2.
Улучшение
качества
воздуха
Учитывая
вклад
деревьев в
улучшение
окружающей
среды,
создание
благоприятных
мест для
отдыха и их
роль в
качестве
поставщика
сырья –
древесины,
они имеют
общепризнанное
значение. При
этом нужно
отметить,
что
способность
к выделению
кислорода
существенно
зависит от
типа
деревьев.
Сравнение
активности
фотосинтеза
некоторых
лиственных
(рис. 1) и
хвойных (рис. 2)
деревьев
дает возможность
предложить
проведение
мероприятий,
способствующих
значительному
улучшению
качества воздуха
в больших
городах
(разумеется,
при этом
нужно
принимать во
внимание и
другие аспекты
промышленного
развития
населенных пунктов).
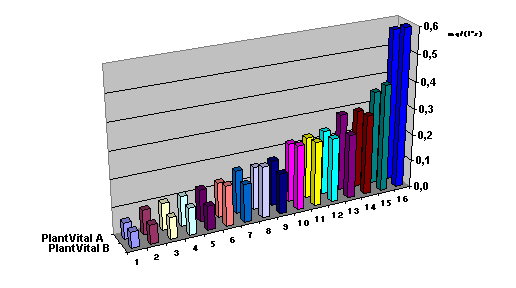
ис. 1.
Сравнение
активности
фотосинтеза
(параметр «S») некоторых
лиственных
деревьев при
одинаковых
условиях
измерения: 1 –
акация, 2
- черёмуха, 3 –
вишня, 4 -
сирень, 5 –
жасмин, 6 – липа, 7
– лигустер, 8 -
смородина, 9 –
каштан, 10 – зеленая
изгородь, 11 –
клен, 12 – калина,
13 – шиповник, 14 –
ива, 15 – тополь, 16 –
береза.
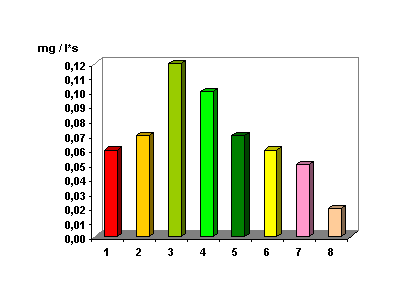
Рис. 2.
Сравнение
активности
фотосинтеза
(параметр «S»)
различных
хвойных
деревьев при
одинаковых
условиях
измерения: 1 –
арча зеленая,
2 – арча голубая,
3 – сосна, 4 – кедр, 5 –
ель, 6 -
ёлка зеленая,
7 – ёлка
голубая, 8 –
лиственница.
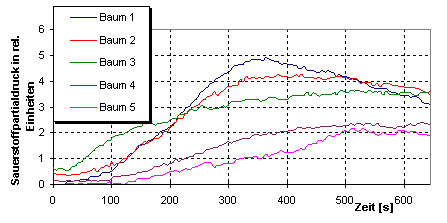
Рис. 3.
Фотосинтетическая
активность
деревьев
липы,
выявленная с
помощью измерительной
системы PlantVital®5000
3.
Повреждения
городских
деревьев в
районах,
подверженных
антропогенному
воздействию
На рис. 3
представлены
измерения
фотосинтетической
активности
деревьев
(липы), произрастающих
на улицах, с
подозрением
на
повреждения.
С помощью
этих
измерений, проведенных
методом
измерительной
системы PlantVital®5000,
были
выявлены, как
только что
начинающееся,
визуально не
видимое
повреждение
деревьев
(дерево 3), так и
ярко
выраженное
негативное
действие
загрязнения
(деревья 4,5). Для
сравнения
служили два
здоровых, не
подверженных
поражению
дерева липы с
одного
проспекта
(деревья 1,2).
Визуальный
осмотр
деревьев
показал
небольшие
изменения у
деревьев 4 и 5, у
дерева 3 не
наблюдалось
изменений.
Обследования
проспекта
показали
небольшие
повреждения
газопровода
под землей, что
послужило
причиной
поражения
деревьев.
После
устранения
этих
повреждений
деревья в
течение двух
лет
вернулись в
свое
нормальное
состояние.
Новый
способ
биоиндикации
является
важным
вкладом для
непрерывного
контроля тренда
нагрузки
опасных
веществ на
местность и
её природные
ресурсы и
может
служить основополагающим
методом для
заблаговременного
проведения
мероприятий
при
превышении допустимой
величины
этой
нагрузки и
связанных с
этим
опасностями
для данной
местности.
4.
Изучение
состояния
кедровых
деревьев в природных
условиях
Совместно с
Институтом
генетики и
разведения
леса
федерального
исследовательского
учреждения
лесного хозяйства
(BFH, Waldsiversdorf), были
проведены
исследования
кедровых культур.
Материал
для
исследования
был выбран из
двух опытных
хозяйств:
лесной школы
Вальдсиверсдорф
и клонового
архивного
хозяйства.
Отбор проб и
производство
анализов
осуществлялось при
одинаковых
условиях.
Детальное
обследование
отдельного
дерева с 4-х
сторон света (С,В,Ю,З),
анализ
фотосинтетической
активности одно- и
двухгодичной
хвои (2002,2003)
показали
незначительный
разброс в данных,
составляющий
всего 8,6 %,
причем
существенного
различия в
активности
фотосинтеза
одно- и
двухгодичной
хвои не
установлено.
Позиция же
веток играет
определенную
роль: хвоя,
расположенная
с южной
стороны,
обладает большей
активностью,
образцы с
других направлений
существенно
не
отличаются.
Сравнение
данных
измерения
активности
фотосинтеза
хвои одного
возраста
выявило
незначительные
колебания
этой
величины.
Процесс изменения
окраски хвои
фенотипичен
и в значительной
мере зависит
от генотипа
растений.
Растения
одного
генопита
(потомство
при
одинаковых
родительских
корнях)
проявляют
себя
достаточно
однородно.
Было
изучено
влияние
пространственного
расположения
хвои на
ветках.
Показано, что
наибольшей
фотосинтетической
активностью
обладали
образцы,
отобранные
из середины
веток,
наименьшей -
на
верхушках.
Молодая хвоя
реагирует,
вероятно,
более резко
на низкие
температуры,
окончания
иголок на
верхушках
веток имели
заметную
желтую окраску.
Снижение
фотосинтетической
активности
хвои на
концах веток
указывает на возрастное
снижение их
физиологической
активности.
Анализ
полученных
результатов
показал существенную
зависимость
фотосинтетической
активности
хвои от
генотипа
растений. Исследуемые
деревья
относились
к различным
потомствам,
т.е. имели
различные родительские
корни, отличаясь
тем самым по
генотипу.
Измерение
фотосинтетической
активности
хвои
индивидуальных
деревьев
внутри одного
потомства
показало
различные
результаты. С
одной
стороны
имелись
потомства, отдельные
деревья
внутри
которых по
измеряемым
параметрам
гомогенны
(незначительные
колебания
данных), что, возможно,
указывает на
их
незначительное
генетическое
различие.
Такие
потомства, как
правило,
имели одни
родительские
корни. Деревья
других
потомств,
например
номер 13 и 14,
напротив,
значительно
отличались
по фотосинтетической
активности
их хвои. Результаты
измерений
имели
большие
отклонения
от средней
величины. В
данном
случае речь
идет о
потомствах,
индивидуальные
деревья
внутри
которых
имели разные
родительские
корни ( к
примеру, одно
материнское
дерево и различные
отцовские),
что,
естественно,
обуславливает
высокую
генетическую
вариабильность
таких
потомств
В ходе
выполнения
измерений
фотосинтетической
активности
хвои
различных
деревьев
была
отмечена
тесная
зависимость
этой величины
от состояния
хвои, от
наличия
желтых
некрозов или
пятен бурых
оттенков в
ней. Хвоя с
указанными
симптомами
выделяла
значительно
меньшее
количество
кислорода.
Причиной
этого
возможно
явилось поражение
хвои тлей в
летний
период (в
случае
некрозов)
Подробный
анализ
данных
показал, что
количество
выделяемого
хвоей
фотосинтетического
кислорода
для
отдельных
деревьев
внутри
чистой
популяции
имеет незначительное
стандартное
отклонение и
может рассматриваться как
характерный
признак чистой
кедровой
популяции.
 http://de.betterplace.org/how_it_works/for_projects
http://de.betterplace.org/how_it_works/for_projects
Рис. 4:
Сравнение
фотосинтетической
активности
хвои
различных
кедровых
деревьев
(номер 13, 14, ..-
обозначение
потомств в
лесной школе
Вальдсиверсдорф).
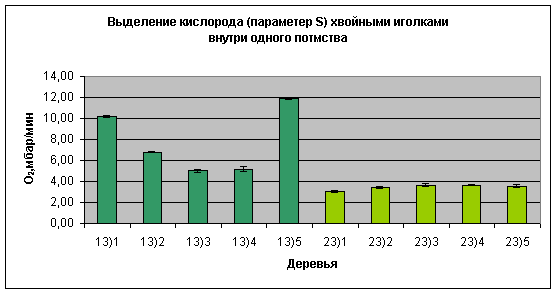
Рис.5:
Сравнение
фотосинтетической
активности
хвои
отдельных
деревьев
внутри одного
потомства
(номер 13, 23 -
обозначение
потомства в
лесной школе
Вальдсиверсдорф,
1,2 .. – номер
дерева
внутри
одного
потомства).
При
исследовании лиственных пород проба растения должна помещаться на мембрану сенсора нижней стороной листа. При
закручивании кюветы следите за тем, чтобы излишек измерительного раствора мог
свободно стекать по сенсору без возникновения давления на мембрану.
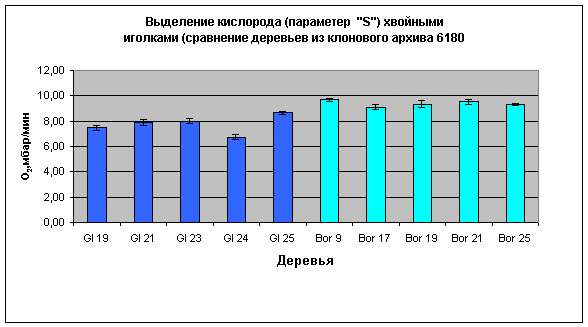
Рис.6:
Сравнение
фотосинтетической
активности
хвои
различных
кедровых
деревьев из архивного
хозяйства (Gl 19.., Bor 9... –
обозначение
деревьев в
клоновом
архивном
хозяйстве).
5.
Исследования
фотосинтетической
активности
листьев
клоновых
культур в
питательной
среде
Совместно с
Институтом
генетики и
разведения
леса
федерального
исследовательского
учреждения
лесного
хозяйства (BFH, Waldsiversdorf),
были
проведены
исследования
на клоновых культурах
некоторых
деревьев.
Первыми
измерениями
фотосинтетической
активности
растительного
материала
изучаемых
культур было
установлено
влияние
продолжительности
культивирования
их в
искусственной
среде на
величину
этой
активности.
Объектом
исследования
служили
клоновые культуры
деревьев,
выращиваемые
в различное
по
продолжительности
время в
специальной
искусственной
питательной
среде (Vitro) :
коричневодревесная
береза (Betula pendula var. carelica), белая
акация (Robinia pseudoacacia), бук (Fagus sylvatica), дуб
черешчатый (Quercus robur). Измерение
фотосинтетической
активности
проводилось
в трех
повторностях
от каждой
клоновой культуры
, объектом
измерения служили
три молодых,
но уже
полностью
развитых
листа. Были
рассчитаны
средние
значения и
стандартные
отклонения
этих трех
измерений.

Рис. 7. Quercus robus Рис. 8. Robinia pseudoacacia
Результаты
проведенных
исследований
для всех
видов
клоновых
культур
показали четкую
зависимость
фотосинтетической
активности
растительного
материала
клоновых
культур от
продолжительности
культивирования
их в
питательной
среде.
Заключение:
Важность
полученных
результатов
состоит в
том, что на основании
достаточно
простых
измерений фотосинтетической
активности
растительного
материала
клоновых
культур в
дальнейшем
возможно с
достаточной
точностью определять
необходимое
время
культивирования
их в
питательной
среде.
Taбл. 1: Пробы
растительного
материала от
клоновых
культур в
питательной
среде
|
№ пп |
Вид
дерева |
Kлон |
Дата
последней смены
питательной
среды |
Замечания |
|
1 |
Дуб
черешчатый |
NL 100A |
25.07.03 |
|
|
2 |
Дуб
черешчатый |
NL 100A |
07.10.03 |
|
|
3 |
Береза
|
2/97 |
15.09.03 |
|
|
4 |
Береза |
2/97 |
29.10.03 |
|
|
5 |
Береза |
BB24 |
16.09.03 |
|
|
6 |
Береза |
BB24 |
29.10.03 |
|
|
7 |
Бук |
Gh12 |
01.08.03 |
|
|
8 |
Бук |
Gh12 |
09.10.03 |
|
|
9 |
Бук |
9193/9 |
08.10.03 |
|
|
10 |
Белая
акация |
2439 |
25.07.03 |
|
|
11 |
Белая
акация |
2439 |
25.09.03 |
|
|
12 |
Белая
акация |
2499 |
03.06.03 |
|
|
13 |
Белая
акация |
2499 |
29.07.03 |
|
|
14 |
Белая
акация |
2444 |
09.10.03 |
Обычные
ростки |
|
15 |
Белая
акация |
2444 |
09.10.03 |
Желатино-подобные
ростки |
|
16 |
Белая
акация |
2498 |
22.10.03 |
Обычные
ростки |
|
17 |
Белая
акация |
2498 |
22.10.03 |
Желатино-подобные
ростки |
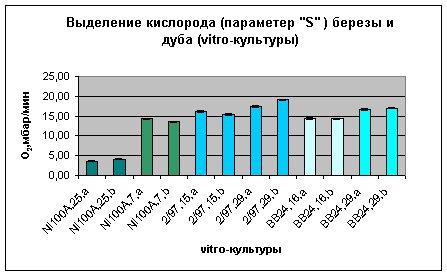
Рис. 9:
Сравнение
фотосинтетической
активности
растительного
материала
различных
клоновых
культур в зависимости
от времени
культивирования
их в
питательной
среде (N1100A.., 2/97.., BB24.. –
обозначение
видов в
клонархиве, a, b –
параллельные
опыты, 25.07... – дата
высадки
культур в
питательную
среду).
В настоящее
время такие
культуры на
основании
многочисленных
литературных
данных, а
также данных
многолетних
собственных
наблюдений
культивируются
в одной
питательной
среде 4-5
недель для
клонов лиственных
деревьев и 6
недель для
клонов
хвойных
деревьев. В
будущем
необходимо
уточнить
оптимальность
установленного
времени
культивации.
6.
Обследование
городской
растительности
в районах,
подверженных
антропогенному
воздействию
С целью
определения
загрязнения
окружающей
среды в областях,
подверженных
антропогенному
воздействию,
были
проведены
измерения
качества
воздуха (см.
соответствующую
руководящую
директиву),
показывающие
фактическое
состояние
воздуха в
момент
измерения. В
отличие от
быстро
изменяющихся
показателей
качества атмосферы
многолетние
растения
(деревья и кустарники)
обладают
способностью
сохранять в
себе
информацию
о вегетационном
развитии с
доминированием
показателей
нового
вегетационного
периода. При
этом
измерение
параметров
жизнеспособности
изучаемых
растений позволяет
оценить
ситуацию
окружающего
региона
более
реально за
длительный
промежуток
времени.
Кроме этого,
здесь может
иметь место
также влияние
вредителей,
из-за которых
в течение
многих лет
страдает
растительность.
На следующих
двух
рисунках
приведены
сравнения
параметра
жизнеспособности
(параметр «S»)
деревьев
липы,
произрастающих
в различных
точках
Берлина и
Штраусберга.
Берлин
характеризуется
наличием
развитого
железнодорожного
узла,
автомобильного
транспорта,
Штраусберг
обладает
курортными
условиями.
Ниже приведены сравнения параметров жизнеспособности деревьев каштана, которые были поражены тлей. Отсюда видно, что сопротивляемость деревьев против вредителей значительно выше в благополучно расположенной местности.
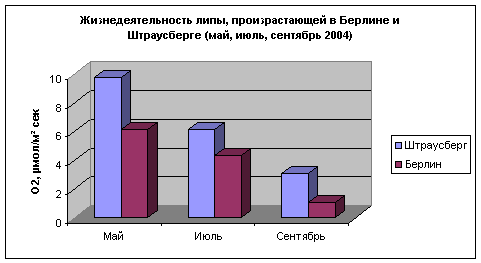
Рис. 10.
Сравнение
значений
параметра
жизнеспособности
«S»
деревьев
липы в
Штраусберге
и Берлине
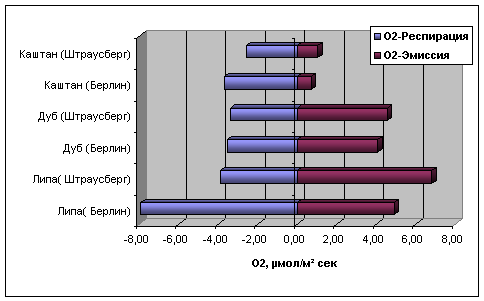
Рис. 11.
Сравнение
значений
параметров
жизнеспособности
«R»
(респирация)
и «S»
(эмиссия)
деревьев
каштана, дуба
и липы в различных
экологических
регионах
(Берлин и Штраусберг)
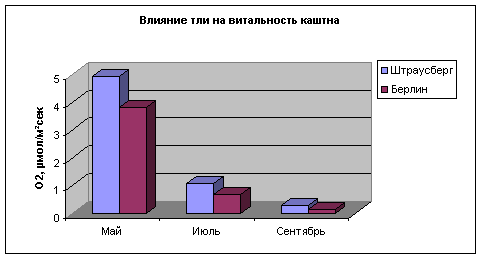
Рис. 12. Сопротивляемость
деревьев
каштана
нападению
тли в
зависимости
от
местонахождения
(Берлин и
Штраусберг)
7.
Сравнительные
измерения
фотосинтетической
активности
некоторых
групп
растений
Сигналы от растений в процессе фотосинтеза можно получить при измерении флоуресценции путем оценки кислородного или углекислого баланса. Все три метода применяются на различных этапах фотосинтетического процесса, поэтому сравнение данных методов представляет большой научный интерес. Учитывая наличие промышленных измерительных систем для потребления углекислоты в процессе фотосинтеза, в таблице указаны литературные данные поглощения СО2. Метод изучения жизнедеятельности растений на основе измерения выделяемого растением кислорода, является сравнительно новым. Поэтому в таблице представлены собственные данные фирмы INNO-Concept GmbH, Штраусберг, полученные для нижеуказанных видов растительности.
Таблица
2: Сравнение
фотосинтетической
активности
некоторых
групп
растений с
помощью
представленных
значений
поглощения СО2 и
эмиссии
кислорода
|
Растение |
Поглощение
СО2 µмол/м²с (литературные
данные) |
Эмиссия
кислорода О2 µмол/м²с (данные
измерений) |
|
С4-растения: |
|
|
|
Кукуруза |
15-54 |
15,7-29,1 |
|
С3-растения: |
|
|
|
Полезные
и
культурные растения |
4,4-35 |
4,4-29,6 |
|
Озимая
пшеница |
10,5-18 |
9,0-12,14 |
|
Лиственные
деревья |
6,0-15,0 |
6,2-15,5 |
|
Хвойные
деревья |
3,0-10,8 |
3,2-8,25 |